TILLING กับการค้นหาสายพันธุ์กลาย |
กนกพร บุญศิริชัย วไลลักษณ์ แพทย์วิบูลย์ วิชัย ภูริปัญญวานิช
กลุ่มวิจัยและพัฒนานิวเคลียร์
สถาบันเทคโนโลยีนิวเคลียร์แห่งชาติ (องค์การมหาชน) |
ในภาษาอังกฤษ คำว่า ‘ till’ เป็นคำกริยา แปลว่า เตรียมดินเพื่อเพาะปลูก เช่น ไถนา ไถกลบ ไถดะ แต่สำหรับ นักอณูพันธุศาสตร์ ‘TILL’ ซึ่งเป็นกริยาเช่นกัน มีความหมายแตกต่างจากการเตรียมดินมากนัก แม้จะไม่ใช่ภาษาเขียน ตามพจนานุกรม ในวงการพันธุศาสตร์เชิงโมเลกุล ‘TILL’ คือ การค้นหาสิ่งมีชีวิตสายพันธุ์กลาย ที่มีการเปลี่ยนแปลง ของลำดับดีเอ็นเอในยีนที่ต้องการด้วยวิธีการ TILLING หรือ Targeting Induced Local Lesions in Genomes ผู้อ่านอาจจะสงสัยว่า ทำไมวิธีการนี้จึงมีชื่อที่ยาวนัก ผู้เขียนเองก็ไม่ทราบคำตอบ เพียงแต่สันนิษฐานว่า เนื่องจาก วิธีการนี้เริ่มใช้กับการศึกษาวิจัยด้านพืชก่อนที่จะมีการนำไปใช้กับสิ่งมีชีวิตอื่น TILLING จึงเป็นคำ ที่อาจดูแล้ว แหมาะสม เพราะมีความหมายเกี่ยวกับการเพาะปลูกพืช จึงได้ตั้งชื่อดังกล่าวขึ้น TILLING คือ กรรมวิธีการทำให้ได้มา ซึ่งสิ่งมีชีวิตที่มีการเปลี่ยนแปลงในยีนที่ต้องการ ซึ่งการเปลี่ยนแปลงนั้นเกิดจาก การเหนี่ยวนำให้เกิดขึ้นในพันธุกรรม ของสิ่งมีชีวิต TILLING จึงมีองค์ประกอบหลัก ๆ สามส่วนคือ ๑. การชักนำให้เกิด การกลายพันธุ์ในสิ่งมีชีวิต ๒. การค้นหาสิ่งมีชีวิตที่มีการกลายในยีนหรือตำแหน่งโครโมโซมที่ต้องการ ๓. การทดสอบหาลักษณะกลายในสิ่งมีชีวิต ที่ได้มา TILLING จึงจัดได้ว่าเป็นการศึกษาทางพันธุศาสตร์แบบ reverse genetics คือ มุ่งเป้าไปที่ยีนกลายก่อน แล้วจึงศึกษาลักษณะกลาย ซึ่งตรงกันข้ามกับ forward genetics ซึ่งมุ่งเป้าไปที่ลักษณะกลายก่อนแล้วจึงศึกษา การกลายในระดับยีน
การชักนำให้เกิดการกลายพันธุ์หรือกระบวนการกลายพันธุ์ (Mutagenesis)ก่อนจะ ‘TILL’ นักวิจัยต้องสร้าง ประชากรสายพันธุ์กลายให้มีขนาดใหญ่เพียงพอ เพื่อจะมั่นใจได้ว่าในประชากรนี้ จะสามารถพบการกลายได้ในทุก ๆ ยีนของสิ่งมีชีวิตชนิดนั้น และโดยทั่วไป มักต้องการแอลลีลกลาย มากกว่าหนึ่งแอลลีลต่อยีน (แอลลีล (allele) คือ รูปแบบของยีน) การคำนวณขนาดของประชากรที่ต้องการ สามารถทำได้โดยใช้สูตร
N = ln(1 - P)/ln(1 - f)
P คือ ความน่าจะเป็นที่จะพบการกลายในยีนที่ต้องการ f คือสัดส่วนของขนาดยีนต่อขนาดของจีโนม N คือขนาดของ ประชากรที่ต้องใช้ในการคัดเลือกเพื่อให้ได้การกลายในยีนที่ต้องการด้วยความน่าจะเป็น P
สิ่งก่อกลายพันธุ์ที่ใช้กันมากกับ TILLING คือ สารเคมีก่อกลายพันธุ์ เช่น ethyl methanesulfonate ( EMS) และ N-nitroso-N-methylurea (NMU) ในพืชเพาะเมล็ดและในแมลงหวี่ และ ethylnitrosourea (ENU) ในปลา และสัตว์เลี้ยงลูกด้วยนม ในประเทศไทย Rice Gene Discovery Unit ซึ่งเป็นหน่วยปฏิบัติการหนึ่งของ ศูนย์พันธุวิศวกรรมและเทคโนโลยีชีวภาพแห่งชาติ สวทช. มีการประยุกต์ใช้ TILLING เพื่อศึกษายีนในข้าว โดยใช้ประชากรที่ได้จากการชักนำให้เกิดการกลายพันธุ์ด้วยอนุภาคนิวตรอนเร็ว สารเคมีก่อกลายพันธุ์และรังสี เช่น รังสีเอกซ์ รังสีแกมมา และอนุภาคนิวตรอนเร็ว มีความคล้ายคลึงกันคือ เป็นสิ่งก่อกลายพันธุ์ที่ไม่มีความจำเพาะ ต่อยีนใดยีนหนึ่ง และนักวิจัยไม่สามารถระบุได้โดยง่ายว่าสิ่งเหล่านี้ก่อให้เกิดการกลายในยีนใด
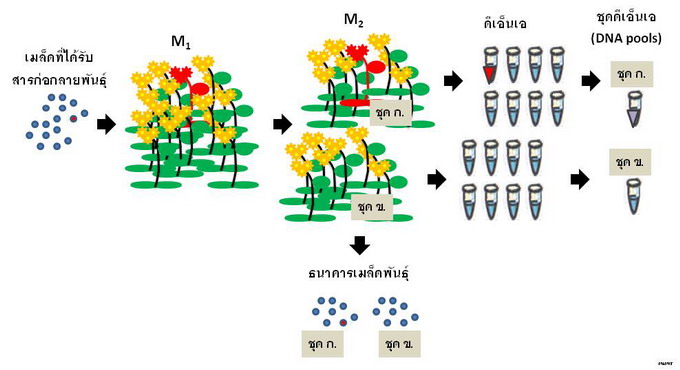
เมื่อชักนำให้เกิดการกลายขึ้นในสิ่งมีชีวิตแล้ว การคัดเลือกลักษณะกลายพันธุ์รวมถึงการประยุกต์ใช้เทคนิค TILLING มักจะไม่ทำในประชากร M 1 (ประชากรที่ได้รับสารก่อกลายพันธุ์โดยตรง) เนื่องจากสารก่อกลายพันธุ์ไม่สามารถส่งผล ให้เกิดการกลายกับยีนเดียวกันในทุก ๆ เซลล์ของสิ่งมีชีวิตได้ สิ่งมีชีวิตในรุ่น M 1 จึงมีลักษณะเป็นไคมีรา (chimera) หรือโมเสก (mosaic) คือแต่ละส่วนของร่างกายอาจมีลักษณะทางดีเอ็นเอที่แตกต่างกัน และลักษณะเหล่านี้ โดยส่วนใหญ่ไม่สามารถถ่ายทอดไปยังรุ่นลูกหลานได้ TILLING จึงอาศัยประชากร M 2 คือรุ่นลูกของประชากร ที่ได้รับสารก่อกลายพันธุ์ เนื่องจากสิ่งมีชีวิตในชั่วรุ่นนี้มีความเป็นหนึ่งเดียวของลักษณะทางดีเอ็นเอภายในตัวสูง ในการทำ TILLING จะสกัดดีเอ็นเอจากส่วนของสิ่งมีชีวิตทุกชีวิตในประชากร M 2 เพื่อจะนำดีเอ็นเอนั้นมาคัดเลือก หาการกลายในยีนที่ต้องการ แต่ผู้วิจัยยังจะต้องให้ประชากร M 2 ขยายพันธุ์ต่อไป เพื่อที่จะระบุตัวสิ่งมีชีวิต ที่มีการกลายนั้นในประชากร M 3 หรือในชั่วรุ่น (generation) ถัด ๆ ไป และนำสิ่งมีชีวิตนั้นมาทดสอบหาลักษณะกลาย เพื่อนำไปใช้ประโยชน์ตามความต้องการ
จะเห็นได้ว่า หากจีโนม ( มวลสาร หรือหน่วย พันธุกรรมทั้งหมดที่จำเป็นต่อการดำรงชีวิตอย่างปกติ ) ของสิ่งมีชีวิต มีขนาดใหญ่ ประชากรที่จะใช้ทำ TILLING ต้องมีขนาดใหญ่ และสำหรับทุกตัวอย่างจะต้องมีการจับคู่ดีเอ็นเอที่ได้ จากชั่วรุ่น M 2 รวมถึงข้อมูลที่ได้จากดีเอ็นเอนั้น กับลูก ๆ ในรุ่น M 3 อย่างเป็นระบบ การวางแผนการจัดการจึงมี ความสำคัญต่อประสิทธิภาพและความสำเร็จของวิธีการนี้เป็นอย่างยิ่ง
ดีเอ็นเอส่วนหนึ่งที่สกัดได้จาก M 2 แต่ละตัวอย่างจะถูกรวมเข้าด้วยกันเป็นชุด ๆ โดยแต่ละชุดจะได้รับดีเอ็นเอ จากตัวอย่างจำนวนเท่า ๆ กัน เช่น ชุด ก. มีดีเอ็นเอจากตัวอย่างที่ ๑- ๘ ชุด ข. มีดีเอ็นเอจากตัวอย่างที่ ๙- ๑๖ รวมชุดละ ๘ ตัวอย่าง ทั้งนี้เพื่อลดงาน ลดระยะเวลา และลดจำนวนปฏิกิริยาลูกโซ่พอลิเมอเรส (polymerase chain reaction หรือ PCR) ที่จะต้องใช้เพื่อการคัดเลือกในเบื้องต้น ดีเอ็นเอส่วนที่เหลือจะถูกเก็บแยกไว้ต่างหากจากกัน เพื่อใช้สำหรับการคัดเลือกในรอบถัดไป |
|
การจัดเก็บดีเอ็นเอและเมล็ดพันธุ์สำหรับพืชผสมตัวเอง ประชากรลูกชั่วรุ่นที่สองจากการได้รับ สารก่อกลายพันธุ์ (ประชากร M 2 ) จะถูกแบ่งออกเป็นชุด ๆ ในที่นี้มีชุดละแปดต้น ดีเอ็นเอจะถูกสกัด จากใบของ M 2 แต่ละต้น แล้วส่วนหนึ่งจากแต่ละตัวอย่างจะถูกรวมกันเข้าเป็นชุดเพื่อนำไปทำ PCR ส่วนเมล็ดที่ได้จากต้น M 2 จะถูกเก็บรวมกันเข้าเป็นชุดไว้ในธนาคารเมล็ดพันธุ์ เพื่อรอการนำไป ศึกษาต่อ |
|
การค้นหาสิ่งมีชีวิตที่มีการกลายในยีนหรือตำแหน่งโครโมโซมที่ต้องการ
โดยที่ TILLING เป็นวิธีการค้นหาการกลายแบบเจาะจงยีน ผู้วิจัยจำเป็นต้องทราบข้อมูลลำดับดีเอ็นเอสำหรับยีนนั้น ๆ หากไม่ทราบจะไม่สามารถค้นหาแอลลีลกลายด้วยวิธีนี้ เนื่องจากเทคนิคนี้อาศัยการเพิ่มจำนวนดีเอ็นเอ เฉพาะส่วนของยีนที่ต้องการด้วยปฏิกิริยาลูกโซ่พอลิเมอเรส ซึ่งต้องใช้ข้อมูลลำดับดีเอ็นเอในการสังเคราะห์ ดีเอ็นเอจุดเริ่ม (primer) ซึ่งจำเป็นในการเริ่มต้นปฏิกิริยาและกำหนดจุดเริ่มต้นและจุดสิ้นสุดของส่วนของยีน ที่ต้องการเพิ่มจำนวน ดีเอ็นเอที่ใช้เป็นต้นแบบในการเพิ่มจำนวนคือดีเอ็นเอที่รวบรวมไว้เป็นชุด ๆ ที่ได้กล่าวถึงแล้ว ดังนั้นดีเอ็นเอที่ได้จากแต่ละปฏิกิริยาจะเป็นดีเอ็นเอที่จำลองมาจากสิ่งมีชีวิตหลาย ๆ ตัวอย่าง หากในชุดมีดีเอ็นเอ ที่ได้จากตัวอย่างที่เกิดการกลายพันธุ์ ดีเอ็นเอที่จำลองได้ในปฏิกิริยานั้นจะมีทั้งดีเอ็นเอปกติ (ในสัดส่วนที่มากกว่า) และดีเอ็นเอที่มีลำดับเบสเปลี่ยนไป (ในสัดส่วนที่น้อยกว่า)
เครื่องมือที่ใช้ในการหาแอลลีลกลายในปัจจุบันมีสองระบบคือ ๑. การใช้ dHPLC (denaturing high pressure liquid chromotography) และ ๒. การย่อยด้วยเอนไซม์ CEL I nuclease ซึ่งได้จากผักชีฝรั่ง (celery) แล้ววิเคราะห์แยกขนาด ดีเอ็นเอด้วยระบบ LI-COR Gel
dHPLC
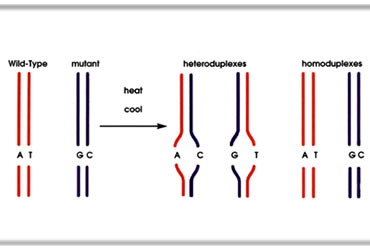
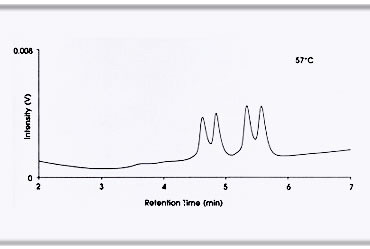
ในการวิเคราะห์หาแอลลีลกลายด้วย dHPLC ดีเอ็นเอที่ได้จากปฏิกิริยาลูกโซ่พอลิเมอเรสจะถูกนำมาทำให้เสียสภาพ ด้วยความร้อน ดีเอ็นเอซึ่งปกติอยู่เป็นเกลียวคู่จะคลายเกลียวแยกออกจากกัน จากนั้นจึงลดอุณภูมิลงเพื่อให้ ดีเอ็นเอคืนสภาพ แต่ดีเอ็นเออาจจะไม่ได้จับเกลียวกับดีเอ็นเอคู่เดิมก็ได้ ดังนั้นในชุดดีเอ็นเอที่มีแอลลีลกลาย จะมีดีเอ็นเออย่างน้อยสามชนิดคือ ๑. homoduplex ของแอลลีลปกติหรือแอลลีลเดิม ๒. homoduplex ของแอลลีลกลาย และ ๓. heteroduplex ระหว่างแอลลีลกลายกับแอลลีลเดิม (homoduplex คือการเข้าคู่กัน ของสายดีเอ็นเอจากแอลลีลเดียวกัน และ heteroduplex คือการเข้าคู่กันของสายดีเอ็นเอจากแอลลีลที่ต่างกัน) หากไม่มีแอลลีลกลาย จะพบ duplex เพียงชนิดเดียวคือ homoduplex ของแอลลีลเดิม เมื่อนำตัวอย่างดีเอ็นเอไปผ่าน คอลัมน์ dHPLC ดีเอ็นเอ duplex เหล่านี้จะเคลื่อนที่ผ่านคอลัมน์ด้วยความเร็วแตกต่างกัน ปริมาณและจำนวนชนิด ของดีเอ็นเอ duplex สามารถประเมินได้จากโครมาโทแกรม (chromatogram) โดยดูจากจำนวนพีก (peak) ของค่าการดูดกลืนแสงที่ความยาวคลื่น 260 nm ของดีเอ็นเอที่ผ่านออกมาจากคอลัมน์เปรียบเทียบกับผล dHPLC ของดีเอ็นจากสิ่งมีชีวิตต้นแบบ จึงสามารถบ่งบอกได้ว่ามีแอลลีลกลายในชุดตัวอย่างหรือไม่ เมื่อพบว่าในชุดตัวอย่าง มีแอลลีลกลายจึงตรวจสอบตัวอย่างดีเอ็นเอจากสิ่งมีชีวิตในชุดนั้นเป็นรายตัว เมื่อพบตัวอย่างที่มีการกลายเกิดขึ้นแล้ว จึงนำลูกหลานของตัวอย่างนั้นไปตรวจสอบศึกษาลักษณะกลายและตำแหน่งของการกลายต่อไป |
|
การเกิด heteroduplex เมื่อดีเอ็นเอที่ได้จากปฏิกิริยาลูกโซ่พอลิเมอเรสประกอบด้วยแอลลีลปกติ (wild-type) และแอลลีลกลาย (mutant) ได้รับความร้อน เกลียวคู่ของดีเอ็นเอจะคลายออกจากกัน และเมื่ออุณหภูมิลดลง สายดีเอ็นเอจะมาเข้าคู่กันใหม่ โมเลกุลที่ได้อาจประกอบด้วยแอลลีลปกต ิจับกับแอลลีลปกติ แอลลีลกลายกับแอลลีลกลาย (สองชนิดนี้จัดเป็น homoduplex ) หรือ แอลลีลปกติ จับกับแอลลีลกลาย (heteroduplex) (ภาพ: Migal-Galilee Technology Center) |
|
ผลจาก dHPLC heteroduplex แสดง retention time ที่แตกต่างจาก homoduplex เมื่อนำมาแยกด้วยเทคนิค dHPLC ที่อุณหภูมิปานกลางซึ่งทำให้ดีเอ็นเอคลายเกลียวเพียงบางส่วน (partial denaturation ) (ภาพ: Migal-Galilee Technology Center) |
|
LI-COR Gel System
สำหรับ LI-COR System ดีเอ็นเอจุดเริ่มทั้งสองชิ้นที่ใช้ในปฏิกิริยาลูกโซ่พอลิเมอเรส จะต้องติดฉลากด้วยสารเรืองแสง (สารฟลูออเรสเซนซ์) ซึ่งให้แสงที่มีความยาวคลื่นแตกต่างกัน หลังจากทำปฏิกิริยาลูกโซ่พอลิเมอเรส ดีเอ็นเอที่ได้ จะถูกทำให้เสียสภาพด้วยความร้อน แล้วคืนสภาพด้วยการลดอุณภูมิ จากนั้นดีเอ็นเอจะถูกย่อยด้วยเอนไซม์ CEL I nuclease เอนไซม์นี้จะตัดสายดีเอ็นเอของ heteroduplex หนึ่งสายที่ตำแหน่งกลาย
เมื่อนำมาแยกขนาดด้วย denaturing gel electrophoresis ในระบบ LI-COR gel ดีเอ็นเอเกลียวคู่จะถูกแยกออกจากกัน
โดยผู้วิจัยสามารถ ตรวจพบแถบดีเอ็นเอได้จากการเรืองแสงของฉลากที่ติดไว้กับดีเอ็นเอจุดเริ่ม
เมื่ออ่านค่าจากการเรืองแสงที่แต่ละ ความยาวคลื่น
ชุดตัวอย่างที่ไม่มีแอลลีลกลายจะพบแถบดีเอ็นเอเพียงหนึ่งแถบซึ่งมีน้ำหนักโมเลกุล
หรือความยาว ของสายดีเอ็นเอใกล้เคียงกันสำหรับทั้งสองความยาวคลื่น
ในชุดตัวอย่างที่มีแอลลีลกลายจะตรวจพบแถบดีเอ็นเอ เพิ่มอีกหนึ่งแถบซึ่งมีความยาวของสายดีเอ็นเอที่
แตกต่างกันในแต่ละความยาวคลื่น และมีค่าน้อยกว่าความยาว ของสายดีเอ็นเอจากแอลลีลปกติ
เมื่อบวกความยาวของสายดีเอ็นเอจากแอลลีลกลายที่ทั้งสองความยาวคลื่น เข้าด้วยกัน จะต้องมีค่าใกล้เคียงกับความยาวของสายดีเอ็นเอจากแอลลีลปกติ
จึงจะสามารถยีนยันได้ว่ามีแอลลีลกลาย ในชุดตัวอย่างนั้นจริง เมื่อพบว่าในชุดตัวอย่างมีแอลลีลกลายจึงตรวจสอบตัวอย่างดีเอ็นเอจากสิ่งมีชีวิต ในชุดนั้นเป็นรายตัว เพื่อระบุตัวสิ่งมีชีวิตที่มีการกลายตามที่ตรวจพบ |
|
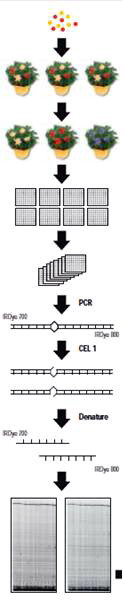
๑. ชักนำให้เกิดการกลายในเมล็ดแล้วนำไปเพาะ
๒. ให้ประชากร M 1 ผสมตัวเอง นำเมล็ดที่ได้ไปเพาะ
๓. ให้ต้น M 2 ผสมตัวเอง นำเมล็ด ที่ได้เก็บไว้ใน ธนาคารเมล็ดพันธุ์ และเก็บใบจากต้น M 2 รายต้น ไปสกัดดีเอ็นเอ
๔. เก็บดีเอ็นเอในถาดหลุมเก็บตัวอย่าง ๑ ตัวอย่างต่อ ๑ หลุม
๕. นำดีเอ็นเอส่วนหนึ่งจากแต่ละหลุมมารวมใส่ถาดใหม่ โดยนำดีเอ็นเอจาก ๘ ถาดในข้อ ๔ มารวมกันเป็น ๑ ถาดชุด
๖. ทำปฏิกิริยา PCR โดยใช้ไพรเมอร์หรือดีเอ็นเอจุดเริ่ม ที่ติดฉลากด้วย IRDye TM 700 และ IRDye TM 800 จากนั้นนำดีเอ็นเอที่ได้มาให้ความร้อนแล้วปล่อยให้เย็นลง ก่อให้เกิด heteroduplex ระหว่างแอลลีลปกติกับแอลลีลกลาย
๗. ตัดดีเอ็นเอ duplex ที่ได้ด้วยเอนไซม์ CEL I nuclease
๘. ดีเอ็นเอถูกทำให้เสียสภาพแล้วนำมาแยกขนาดด้วยวิธี electrophoresis โดย LI-COR 4300 DNA Analysis System
๙. ได้ภาพของ LICOR gel จากสองช่องสัญญาณ IRDye TM 700 และ IRDye TM 800
( ภาพ: LI-COR Inc. 2003. Automated Reverse Genetics. www.licor.com)
|
|
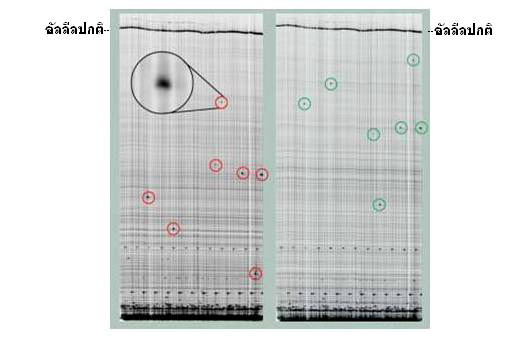
LI-COR Gel รูปจากช่องสัญญาณ IRDye TM 700 (ซ้าย) และ ช่องสัญญาณ IRDye TM 800 (ขวา) เมื่อ heteroduplex ถูกตัดด้วย CEL I ที่ตำแหน่งกลาย ผลรวมคือ ในแต่ละตัวอย่างจะได้ ดีเอ็นเอที่ติดฉลากด้วย IRDye TM 700 (วงกลมสีแดง) และ IRDye TM 800 (วงกลมสีเขียว) ที่มีขนาดสั้นกว่าแอลลีลปกติ แต่เมื่อขนาดของทั้งสองรวมกันแล้วจะมีค่าเท่ากับขนาดของแอลลีลปกติ ( ภาพ: LI-COR Inc. 2003. Automated Reverse Genetics. www.licor.com)
|
|
การทดสอบหาลักษณะกลาย
เมื่อได้สิ่งมีชีวิตที่มีแอลลีลกลายที่ต้องการในสภาพฮอโมไซกัส ( homozygous) คือที่ตำแหน่งของยีนนี้บนโครโมโซม ทุกชุด เป็นแอลลีลกลายเดียวกันทั้งหมด โดยไม่มีแอลลีลเดิมบนโครโมโซมชุดใด ๆ จึงนำสิ่งมีชีวิตนั้นมาทดสอบ ลักษณะกลาย เนื่องจากการทำ TILLING เป็นการเจาะจงศึกษายีนใดยีนหนึ่ง ผู้วิจัยจึงมักมีสมมติฐานอยู่แล้วว่ายีนนี้ ทำงานเกี่ยวข้องกับระบบใดของสิ่งมีชีวิต เมื่อได้สิ่งมีชีวิตสายพันธุ์กลายมาจึงสามารถเจาะจงทดสอบการทำงาน ของระบบนั้น ๆ ได้ เช่น หากเรา ‘TILL’ ในยีน DWARF ซึ่งเราทราบว่าควบคุมความสูงของพืช เราก็คาดหวังว่า พืชสายพันธุ์กลายที่ได้จะเตี้ยกว่าปกติ สิ่งที่ต้องทำคือปลูกพืชแล้วเปรียบเทียบความสูง หาก ‘TILL’ ในยีนที่ผลิตเอนไซม์ catalase เราก็ต้องตรวจหา catalase activity
ในบางกรณี เราอาจตรวจไม่พบลักษณะกลายใด ๆ เลย เมื่อเกิดกรณีเช่นนี้ขึ้น ผู้วิจัยจำเป็นต้องตรวจดูลำดับดีเอ็นเอ ของยีนกลายว่าการกลายที่เกิดขึ้นเป็น silent mutation หรือไม่ silent mutation คือการกลายที่ไม่ก่อให้เกิด การเปลี่ยนแปลงในลำดับกรดอะมิโนของโปรตีนที่เป็นผลลัพธ์จากการถอดรหัสของยีนนั้น silent mutation โดยส่วนมากจะไม่ก่อให้เกิดลักษณะผิดปกติใด ๆ ในสิ่งมีชีวิตเลย
หลายครั้งที่สมมติฐานที่นักวิจัยตั้งไว้เกี่ยวกับลักษณะกลายที่คาดว่าจะได้รับไม่ถูกต้อง โดยเฉพาะในการศึกษายีนที่เป็น paralog ของยีนอื่น (ยีนที่มีลำดับดีเอ็นเอคล้ายกันแต่อาจมีการทำงานที่แตกต่างกัน) ลำดับกรดอะมิโนที่ถอดรหัสได้ และสัณฐานทางโมเลกุลของโปรตีนที่เป็นผลลัพธ์ของยีนนั้น อาจบ่งบอกว่ายีนนี้มีหน้าที่เฉพาะอย่างใดอย่างหนึ่ง ในระดับเซลล์ ซึ่งนักวิจัยมักจะสามารถตั้งสมมติฐานไว้ได้อย่างถูกต้อง แต่ในระดับสิ่งมีชีวิต ยีนนี้อาจทำงานในเนื้อเยื่อ ที่จำเพาะและในช่วงเวลาที่จำเพาะมาก ๆ โดยไม่ทำงานในเนื้อเยื่ออื่นหรือในเวลาอื่นเลย ซึ่งในส่วนนี้นักวิจัยอาจจะ ไม่มีข้อมูลอยู่ สมมติฐานเกี่ยวกับลักษณะกลายที่ควรพบจึงมีโอกาสผิดพลาดได้ ดังนั้นหากผู้วิจัยตรวจไม่พบ ลักษณะกลายในระบบที่ตั้งสมมติฐานไว้ อาจจำเป็นต้องศึกษาหาข้อมูลเกี่ยวกับการทำงานของยีนนั้น ๆ โดยละเอียด ก่อนที่จะออกแบบการทดลองเพื่อตรวจหาลักษณะกลายอีกครั้งหนึ่ง
|
บริการ TILLING ในปัจจุบัน
TILLING เป็นเทคนิคที่ช่วยให้นักวิจัยค้นหาสิ่งมีชีวิตที่มีการกลายในยีนที่ต้องการได้ และอาจสามารถทำได้โดยที่ไม่ต้องการการลงทุนในส่วนของเครื่องมือมากนัก แต่ต้องมีการลงทุนในการจัดระบบการเก็บและวิเคราะห์ตัวอย่าง พื้นที่ และแรงงานค่อนข้างมาก และจำเป็นต้องเก็บตัวอย่างจำนวนมากไว้ตลอดระยะดำเนินการของโครงการ ดังนั้นโครงการ TILLING โดยส่วนใหญ่จึงไม่ใช่การทำขึ้นเพื่อใช้เองภายในห้องปฏิบัติการห้องใดห้องหนึ่ง แต่เปิดให้บริการแก่ห้องปฏิบัติการอื่นทั่วประเทศหรืออาจจะทั่วโลกที่มีความสนใจศึกษาสิ่งมีชีวิตชนิดเดียวกัน ปัจจุบันมีโครงการ TILLING ที่เปิดให้บริการมากมาย ดังตัวอย่างที่แสดงไว้ในตาราง |
หน่วยงานที่ให้บริการ |
สิ่งมีชีวิตที่เปิดบริการ |
สถานที่ |
เว็บไซต์ |
Seattle TILIING Project |
Arabidopsis thaliana
Drosophila melanogaster |
Seattle , WA, USA |
tilling.fhcrc.org |
UC Davis Genome Center |
Oryza sativa |
University of California, Davis, CA, USA |
tilling.ucdavis.edu |
CAN-TILL |
Arabidopsis thaliana
Caenohabditis elegans
Brassica oleracea |
University of British Columbia, Vancouver, B.C., Canada |
www.botany.ubc.ca/can-till/ |
The Maize TILLING Project |
Zea mays |
USA |
genome.purdue.edu/maizetilling/index.htm |
RevGenUK |
Lotus japonicas
Medicago truncatula |
UK |
revgenuk.jic.ac.uk |
Zebrafish TILLING Project |
|
USA |
webapps.fhcrc.org/science/tilling/index.php |
|
เอกสารอ้างอิงและอ่านเพิ่มเติม
- Comai, L., and Henikoff, S. 2006. TILLING: practical single-nucleotide mutation discovery. Plant J. 45: 684-694.
- Cooper, J.L., et al. 2008. TILLING to detect induced mutations in soybean. BMC Plant Biol. 8:9-18.
- Kovar, J. et al. 2004. Application of TILLING ?/EcoTILLING To Screen For Small Mutations in Mammals. www.licor.com.
- Lander, E.S., and Waterman, M.S. 1988. Genomic mapping by fingerprinting random clones: a mathematical analysis”, Genomics 2(3): 231-239.
- LI-COR, Inc. 2003. Automated reverse genetics. www.licor.com.
- McCallum CM et al. (2000) Targeting induced local lesions in genomes (TILLING) for plant functional genomics. Plant Physiology 123, 439-442
- Oleykowski CAet al. (1998) Mutation detection using a novel plant endonuclease. Nucleic Acids Research26, 4597-4602
- Rice Gene Discovery Unit, BIOTEC, NSTDA. www.biotec.or.th/biotechnology-en/Person/Unit-Rice.asp.
- Slade AJ et al. (2005) A reverse genetic, non-transgenic approach to wheat crop improvement by TILLING. Nature Biotechnology23, 75-81
- Till, B.J., et al. 2003. Large-Scale Discovery of Induced Point Mutations With High-Throughput TILLING. Genome Research. 13:524-530.
- Transgenomic, Inc. How dHPLC works for mutation detection. www.transgenomic.com.
|
|
| |